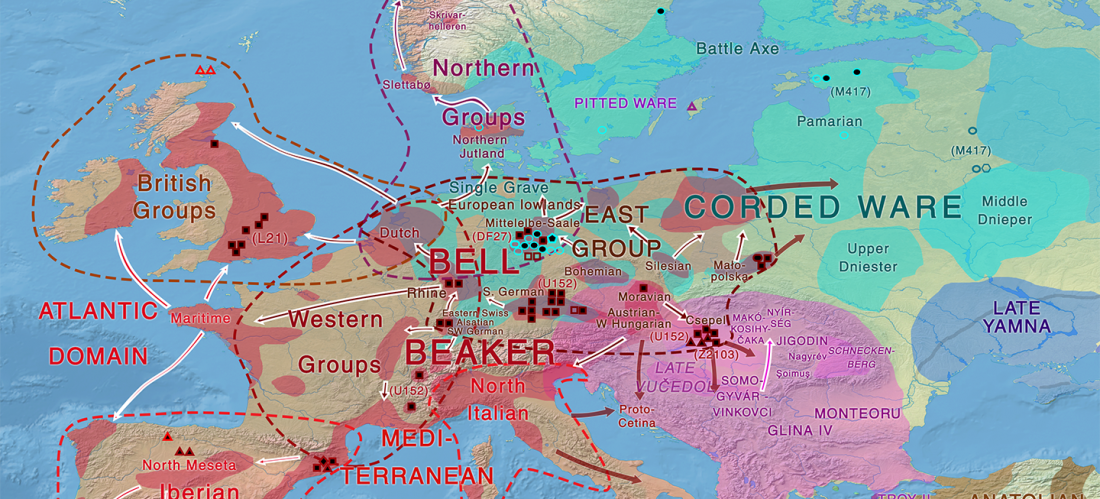
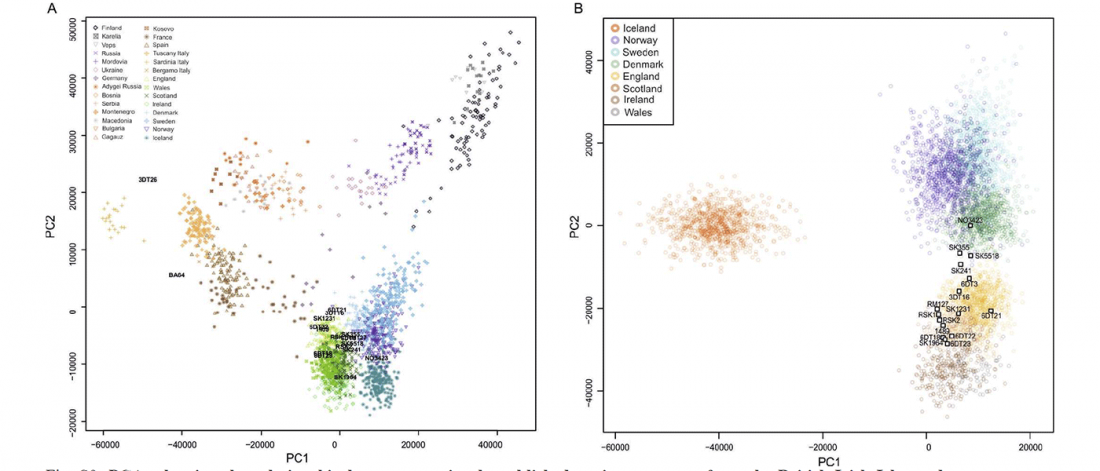
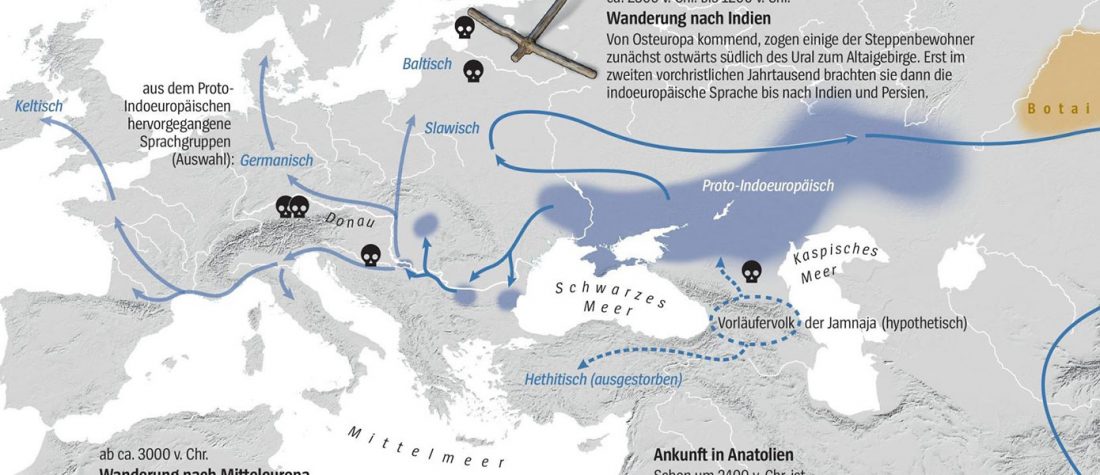
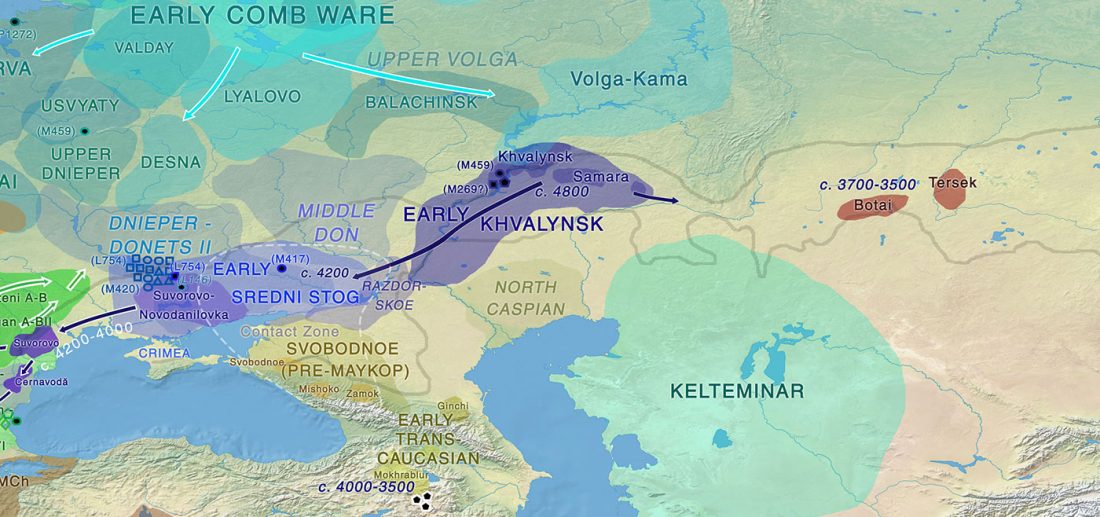
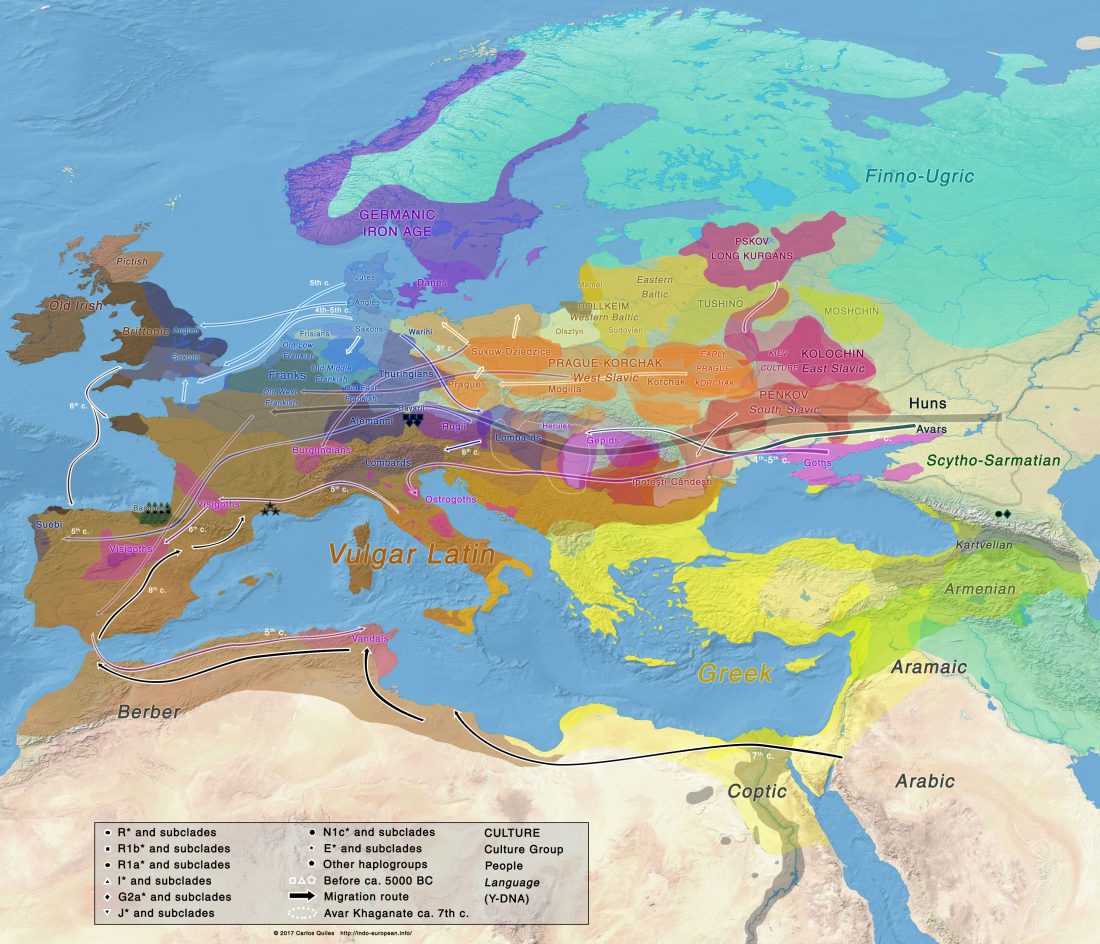
Jamnaja die wahrscheinlichste Quelle für das Hauspferd; die nächste Linie, aus der Glockenbecher-Kultur
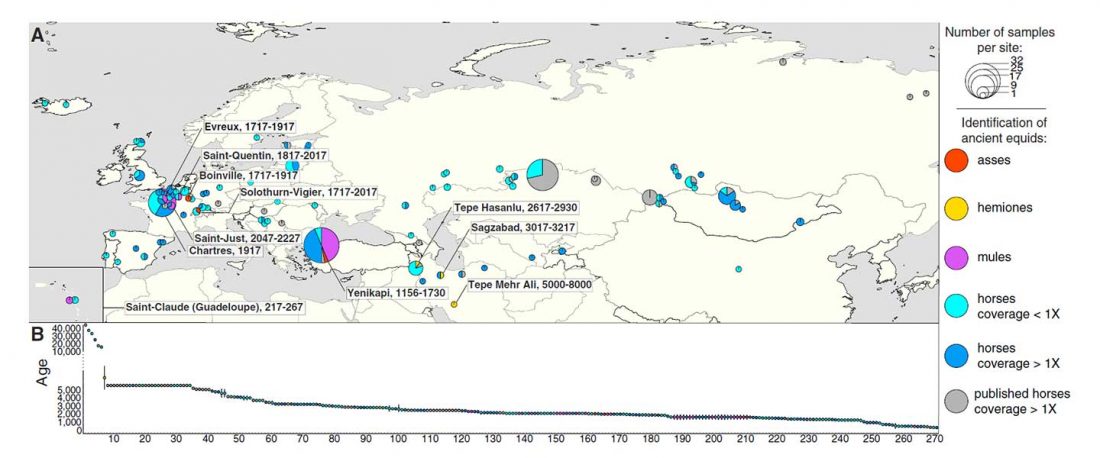
Öffentlich zugängliche Tracking Five Millennia of Horse Management with Extensive Ancient Genome Time Series, von Fages et al. Cell (2019).
Interessante Auszüge:
… Read the rest “Jamnaja die wahrscheinlichste Quelle für das Hauspferd; die nächste Linie, aus der Glockenbecher-Kultur”Der früheste archäologische Beweis für das Melken, Aufspannen und Korrallieren von Pferden findet sich in der ∼5.500 Jahre alten Botai-Kultur der zentralasiatischen Steppen (Gaunitz et al., 2018, Outram et al., 2009; siehe Kosintsev und Kuznetsov, 2013 für Diskussion). Botai-ähnliche Pferde sind jedoch nicht die direkten Vorfahren moderner Domestizierter, sondern der Pferde von Przewalski (Gaunitz et al., 2018). Die genetische Herkunft moderner Domestizierter Pferde ist daher weiterhin umstritten. Vorgeschlagene Kandidaten sind die pontisch-kaspischen Steppen (Anthony, 2007), Anatolien (Arbuckle,