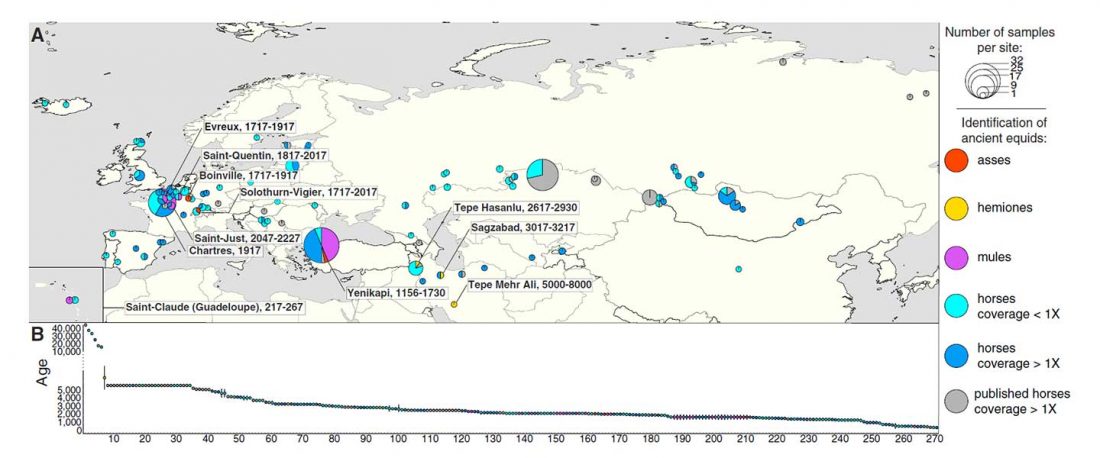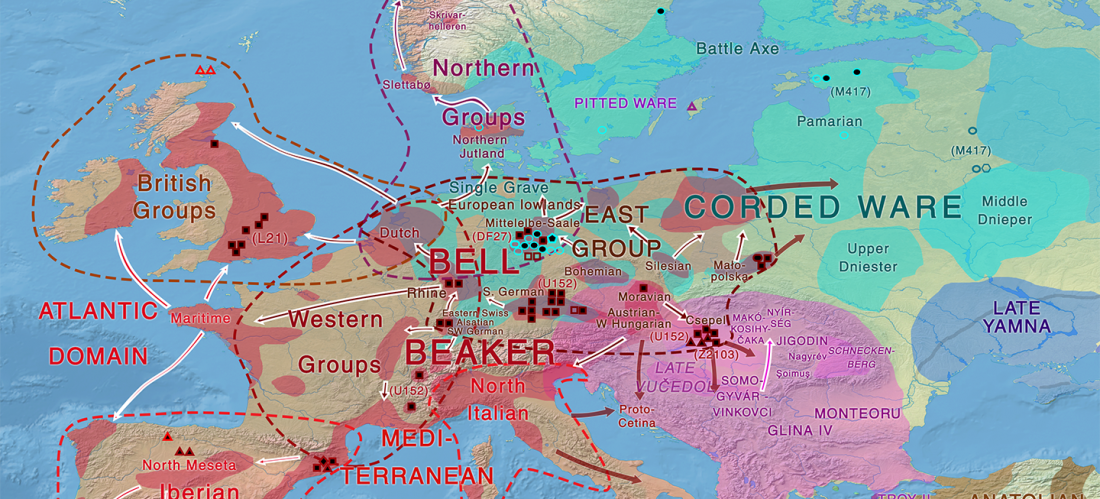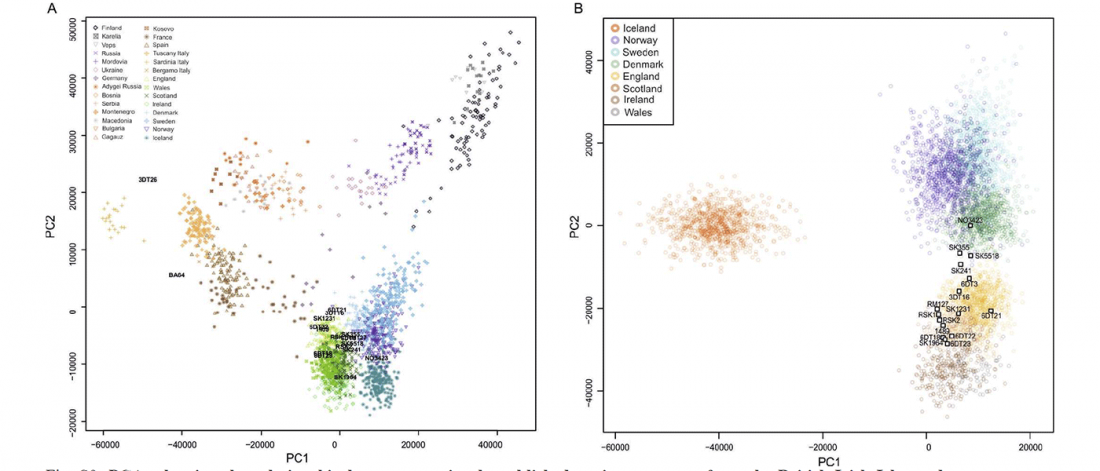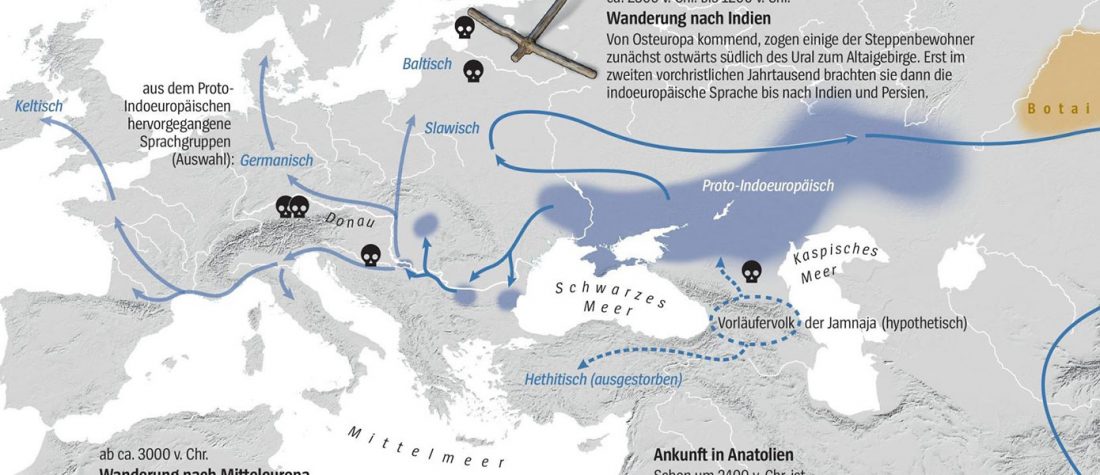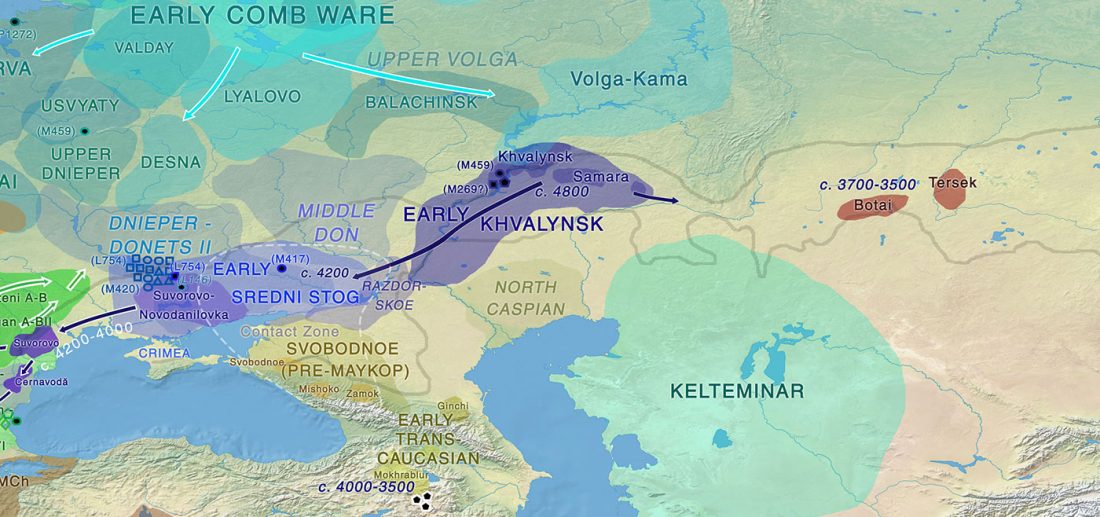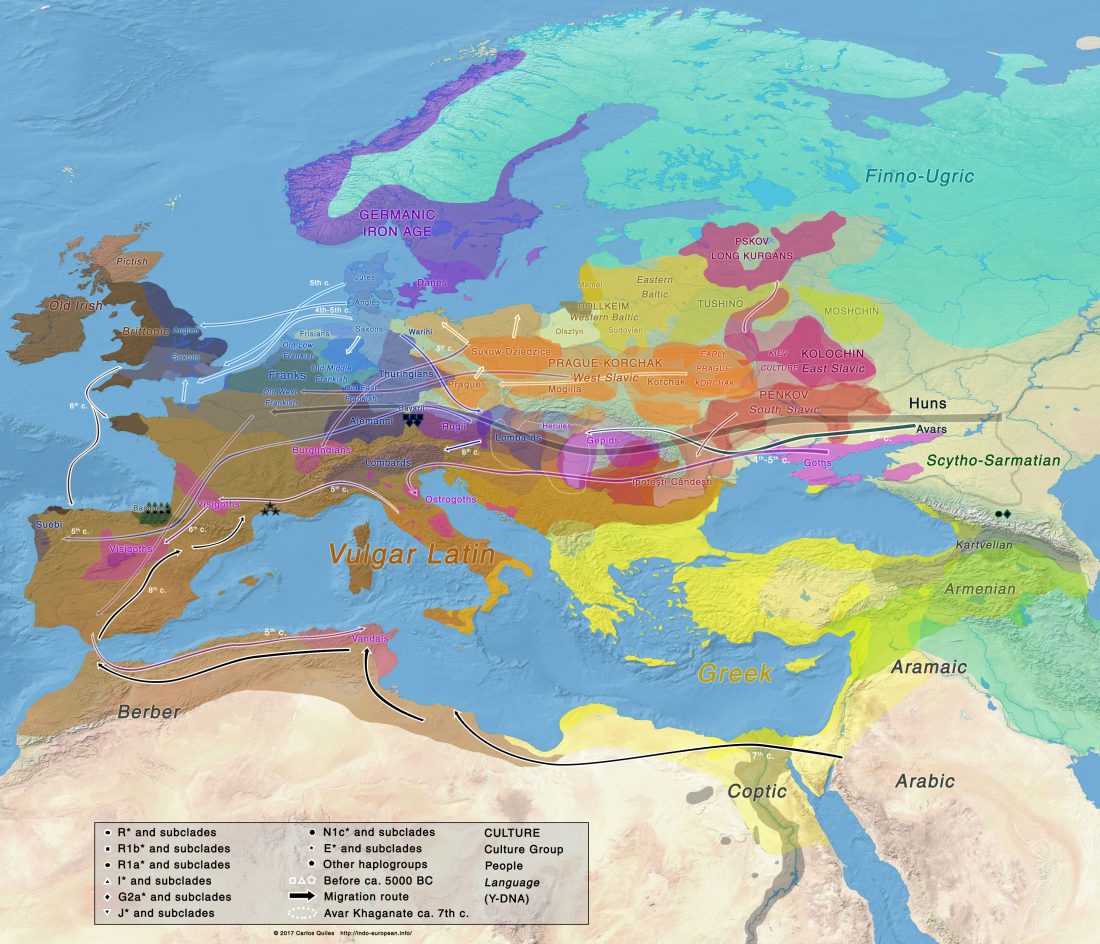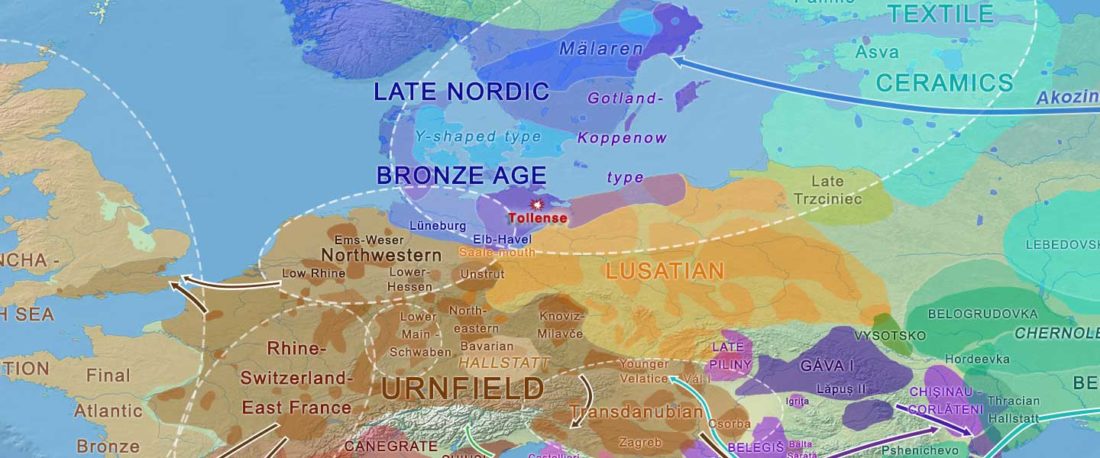
Kürzlich veröffentlichtes Genomische Daten von einem alten europäischen Schlachtfeld weisen auf eine anhaltende starke Selektion einer Genomregion hin, die mit der Laktasepersistenz in den letzten 3.000 Jahren assoziiert ist, von Burger et al., Current Biology (2020).
Interessante Auszüge:
Die Proben vom Tollense-Schlachtfeld zeigen keine Struktur
… Read the rest “Bronzezeit-Krieger im Tollensetal zeigen enge Verbindungen mit Langobarden”Mehrere Beweislinien weisen auf eine geringe oder keine genetische Struktur in der Population hin, aus der die Tollense-Individuen entnommen wurden. Erstens fallen alle Individuen in den Bereich der mittel- und nordeuropäischen Variation, wenn sie auf eine Hauptkomponentenanalyse (PCA) projiziert werden, die an modernen Proben trainiert wurde, und ihre Ausbreitung entspricht der anderer Proben der