Recent papers on France and neighbouring regions, Ancient genome-wide DNA from France highlights the complexity of interactions between Mesolithic hunter-gatherers and Neolithic farmers, by Rivollat et al. Science Advances (2020) 6(22), and Ancient genomes from present-day France unveil 7,000 years of its demographic history, by Brunel et al. PNAS (2020).
Interesting excerpts (emphasis mine):
I. Survival of HG ancestry in Central Europe
From Rivollat et al. (2020):
Here, we present newly typed genome-wide data from 101 individuals from 12 sites from modern-day France and Germany (3 Late Mesolithic and 98 Neolithic, 7000–3000 cal BCE (…)
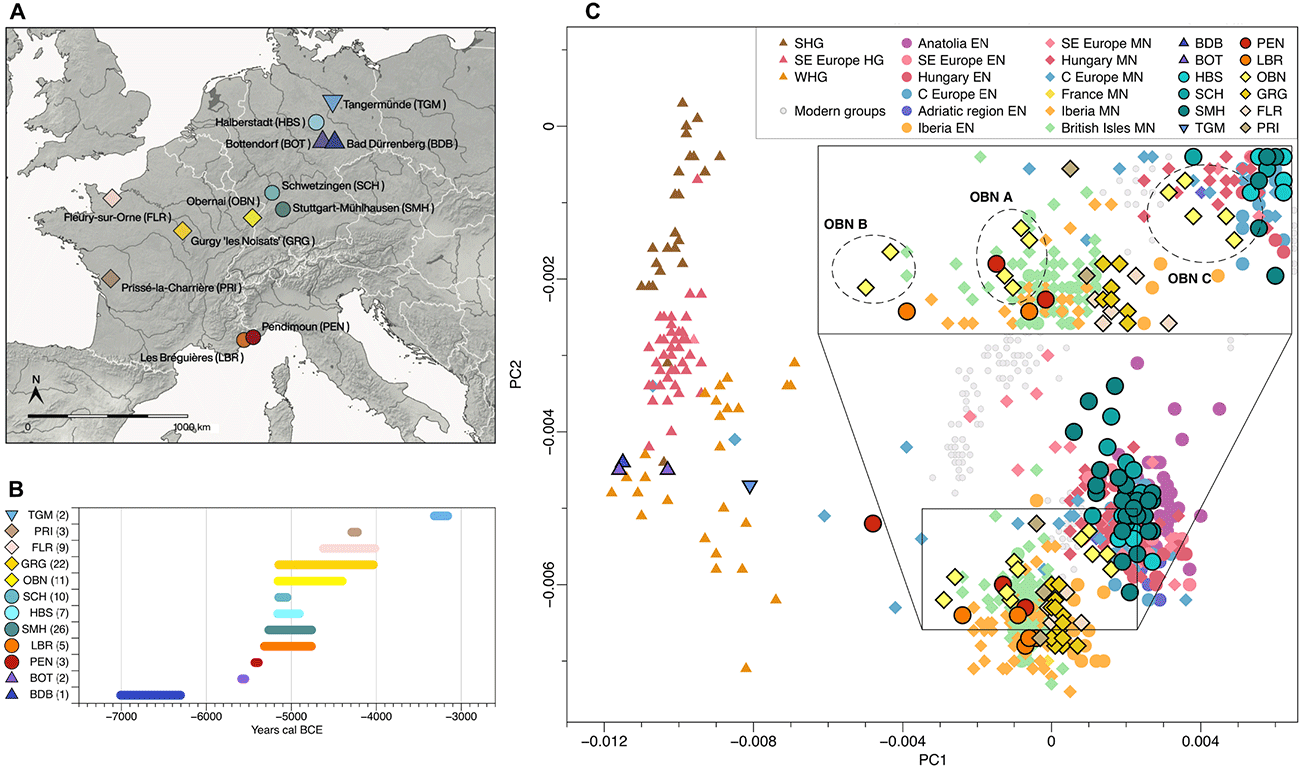
We explored our new data qualitatively using principal components analysis (PCA) by projecting the ancient samples onto the genetic variation of a HO set of west Eurasians. Newly sequenced Late Mesolithic individuals from Bad Dürrenberg (BDB001) and Bottendorf (BOT004 and BOT005) fall within the variability of western HG (WHG). Individual TGM009, attributed to the Late Neolithic site of Tangermünde from an Elb-Havel context [Trichterbecherkultur (TRB)], occupies an intermediate position between WHG and Neolithic farmers. Newly sequenced Neolithic individuals cluster with previously published data, forming two regional subgroups: the first including individuals from Neolithic central and southeastern Europe and the second individuals from Neolithic western Europe (Iberian Peninsula, France, and British Isles), which is shifted slightly toward WHG on PC1. Formal f-statistics of the form f4(Mbuti, European_HG; test, Anatolia_Neolithic), where test represents Neolithic groups, confirm these observations via different degrees of affinity with European HG.
Early Neolithic individuals from Stuttgart-Mühlhausen (SMH), Schwetzingen (SCH), and Halberstadt (HBS) in Germany [44 published and 42 newly reported, of which 5 low-coverage individuals (<18,000 SNPs mapping on the HO panel) are shifted toward the centroid] representing the LBK horizon (17 sites from Hungary, Austria, and Germany) form a homogeneous genetic group of early farmer populations in central Europe. Neolithic groups from France cluster with western Neolithic individuals, while southern French sites Pendimoun (PEN) and Les Bréguières (LBR), attributed to the ICC, are shifted even further toward WHG. This suggests a higher HG component than in any other early Neolithic individuals, and thus a different history of admixture compared to contemporaneous western early farmer groups from Iberia.
Both central and western groups are still present during the fifth and early fourth millennium BCE, suggesting a geographic border along the Rhine River for regions that are situated north of the Alps. Individuals from the Middle Neolithic French sites of Gurgy (GRG), Prissé-la-Charrière (PRI), and Fleury-sur-Orne (FLR) in the northern half of France appear homogeneous (one PRI individual outside of the cloud has low SNP coverage), while the individuals from the site Obernai (OBN) form three groups: one with other contemporary western individuals (OBN A), one with a stronger HG component (OBN B), and a third one with central European farmers (OBN C), despite sharing similar cultural and chronological backgrounds (…)
Origin of HG ancestry
We caution that because the amount of HG ancestry is very low in many Neolithic groups (<10%), it remains difficult to characterize the ultimate source reliably. Nevertheless, our admixture patterns from supported models show clear geographic signals. Neolithic groups associated with the LBK in central Europe (Hungary, Austria, and Germany) carry a small HG proportion, which was likely derived from admixture with HG individuals of the EHG-WHG cline and could have occurred in southeastern Europe during a preceding phase of the Neolithic expansion around 6000–5400 BCE. When using f-statistics of the form f4(Mbuti, test; BDB001, KO1) with our new Mesolithic genome (BDB001) from the Middle-Elbe-Saale region in central Germany as a geographically local HG proxy (instead of using Loschbour, which is located west of the Rhine), we do not find support for a local attraction for LBK groups, but the same pattern as for Loschbour. This suggests that additional gene flow from neighboring Loschbour-like HG such as BDB001 in central Europe was negligible in the first Neolithic groups. However, the German Baalberge group (4000–3500 BCE) shows a marked increase of such HG ancestry, as well as individuals from the Blätterhöhle group, as has been suggested (5, 6), compared to a combination of both KO1-like and Loschbour-like ancestries for LBK groups (6). We can now show that this increase in WHG ancestry (up to 21.3 ± 1.5% in Baalberge) is driven by either local Loschbour-like ancestry or an expansion of farming groups from the west carrying this signal during the fifth millennium BCE, as suggested by archaeological data. For all studied Neolithic groups west of the Rhine, we observe a different pattern with a higher HG ancestry proportion, even for earliest groups that appears to be of a local (Loschbour-like) HG origin, consistent with archaeological data.
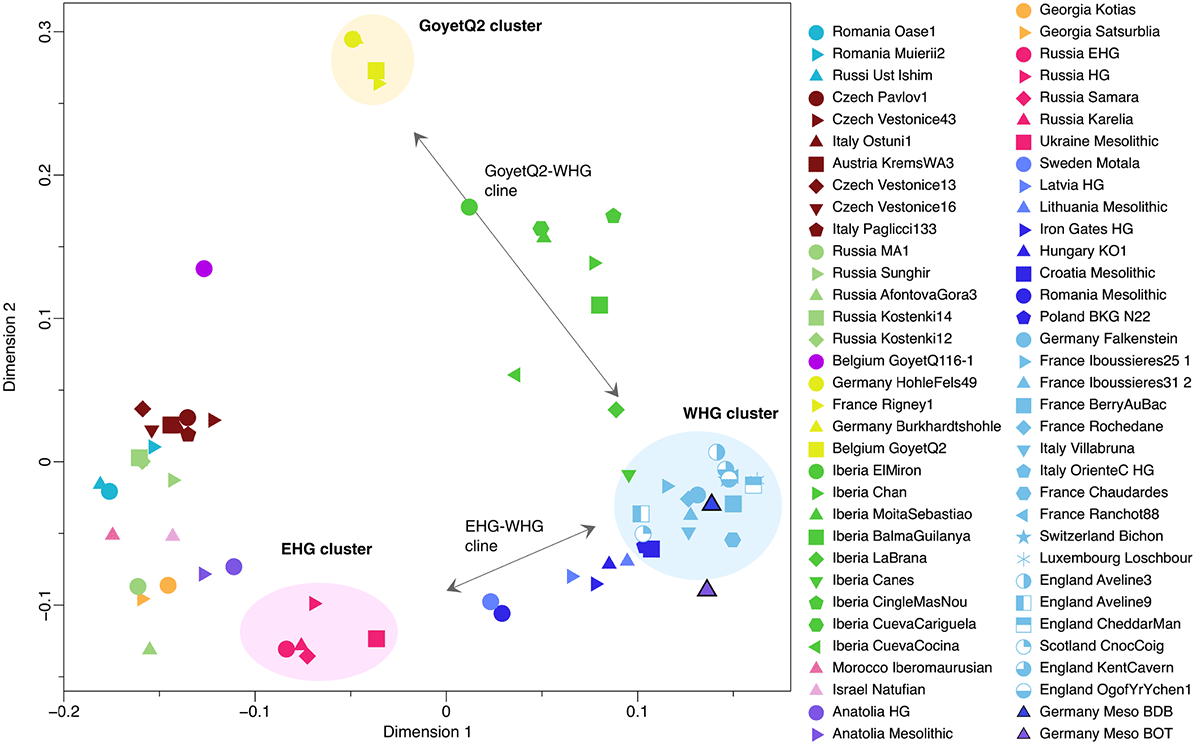
Multidimensional scaling plot of genetic distances based on f-statistics of the form f3(Mbuti; test, test) between Eurasian HG individuals. Newly reported individuals have a black outline.”
Late survival of HG ancestry
In contrast to Middle-German Neolithic groups, the Late Neolithic individual TGM009 (~3300 BCE) found in an TRB/Elb-Havel group context shows a different pattern of HG ancestry. When modeled with distal sources, this individual carries 63.6 ± 5.2% Villabruna-related ancestry. When we use proximal sources (MODEL C; table S13), we estimate the HG component to be split into 48.1 ± 6.4% KO1-related ancestry and 25.8 ± 6.1% Loschbour-like ancestry (P = 0.12). To test whether the subtle EHG-related signal observed in TGM009 (through KO1) could also come from Scandinavian HG, as suggested by the archaeological records of regional late contacts with the Scandinavian Mesolithic (37), we applied f-statistics of the form f4(Mbuti, test; Hungary_KO1, Sweden_Motala_HG). The f4 value is significantly negative (f4 = −0.0035, Z score = −6.383), indicating that TGM009 shares more ancestry with southeastern Europe HG than with Scandinavian HG (table S16). Consequently, qpAdm models with Sweden_Motala_HG in place of Hungary_KO1 received a poor fit (P = 1.32−04; table S16).

II. The arrival of Bell Beakers
From Brunel et al. (2020):
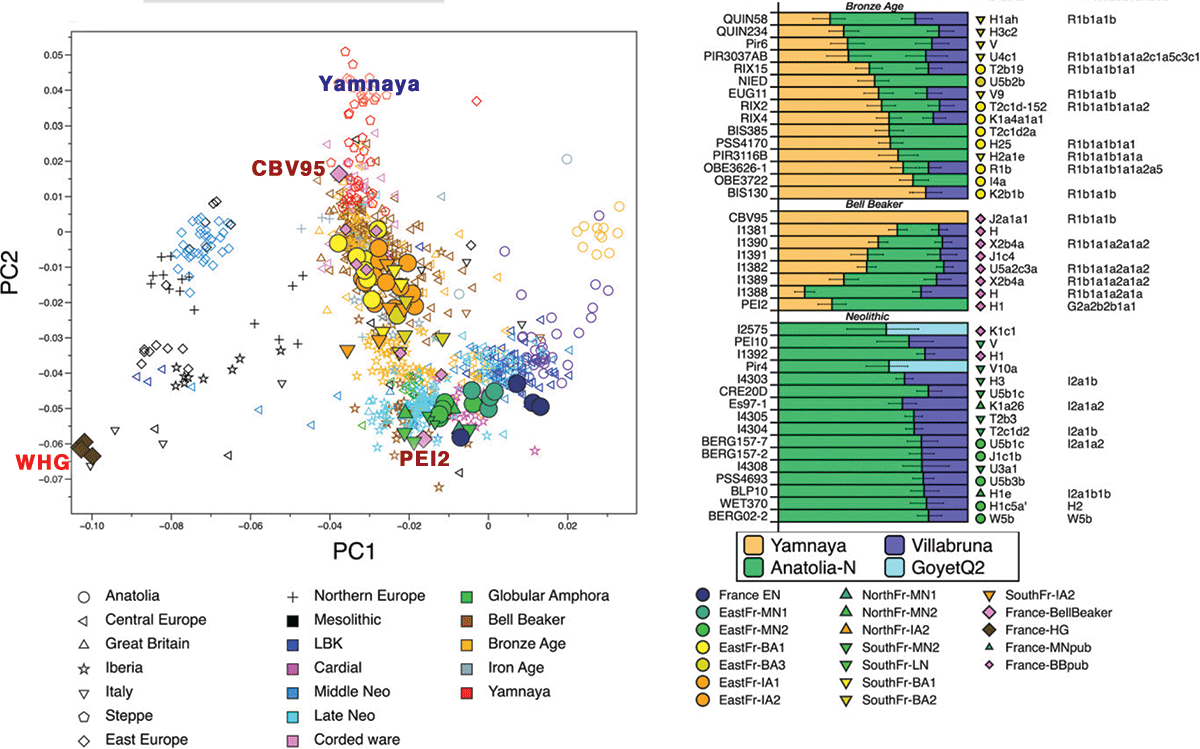
We report two Bell Beaker-associated individuals (CBV95 and PEI2), that we coanalyzed with previously reported contemporaneous individuals from Europe, including France. French Beaker-associated individuals display a wide range of steppe ancestry proportions. CBV95 in northern France derives the highest proportion of alleles from the Yamnaya in our dataset, and belongs to Y-chromosome haplogroup R1b, providing the earliest clear evidence of the presence of this haplogroup in France around 2500 BCE. This lineage was associated with the arrival of migrants from the steppe in central Europe during the Late Neolithic, and was described in other parts of Europe and in Bell Beaker-associated individuals from southern France, while being almost absent in Iberia prior to the Bronze Age (10, 13).
PEI2, a male unearthed from a collective burial site near Carcassonne in southwestern France with artifacts of the Bell Beaker complex, falls within the genetic diversity of Neolithic individuals in the PCA. Modeling admixture. proportions between three source populations, Anatolia_Neolithic, Villabruna, and Yamnaya_Samara, we could, however, detect 28.3% of steppe ancestry in PEI2. These observations are consistent with previous findings and confirm that steppe ancestry appeared later and with a lower impact in southwestern Europe than in other parts of the continent (10, 25). Surprisingly, the admixture model did not include Villabruna as an ancestry source for either CBV95 or PEI2, which differs from previously known Late Neolithic individuals.
Relative Continuity between the Bronze Age and the Iron Age
As foretold by its early occurrence in Beaker-associated CBV95, a drastic Y-chromosome turnover occurs during the Bronze Age, where R1b replaces the preexisting diversity of Neolithic lineages in our sampling (Bronze Age, 11/13 and Iron Age, 7/10 individuals having R1b).
Bronze and Iron Age France share a common space in the PCA plot, both shifted toward modern central Europe and falling within the genetic diversity of Bronze Age Britain and central Europe, with a homogenization of the steppe component. In contrast to what was described for central Europe, there is no further shift toward eastern Eurasian genotypes during the Iron Age.Instead, the steppe component, heterogeneously distributed between individuals during the Bronze Age (ranging from 30 to 70%), becomes homogeneous, and individuals from the Hallstatt and La Tène culture in the French territory display similar affinities toward both modern and ancient populations.
III. Renewed wave of Steppe ancestry
It is an interesting coincidence that the paper on HG survival in West-Central Europe appeared at the same time as the expected paper on the arrival of R1b-rich Bell Beakers from the east, because the former serendipitously complements the findings of the latter.
The main example of this synergy is found in the reported early Bell Beaker sample CBV95, who shows a clear ‘northern’ cluster in the PCA compatible with the incorporation of HG ancestry (despite the reported lack of Villabruna-related admixture, most likely a problem of outgroup selection), a feature this sample has in common with the recently reported Aes25, a Swiss Bell Beaker from the Aesch Dolmen.
NOTE. The likely common Yamnaya+HG-related cluster formed by these samples, with a likely common origin around the Upper Danube region, helps further refine the wide confidence intervals and unlikely early dates reported by Furtwängler et al. (2020), evidently affected by reservoir effects, hence most likely closer to the turn of the 25th century for these HG+Steppe-rich individuals.
While the genotype data hasn’t been released yet, there are indirect ways to compare how this sample will look like. First, its PCA cluster and thus admixture are likely to be similar to the Swiss Aes25 individual, which is compatible with the expected northward migration of East Bell Beakers from the Upper Rhine.
To be able to compare potential origins of their common Steppe-related ancestry, I selected the closest available HG-rich population of the Neolithic, which – barring the publication of samples from the Upper Rhine or the Upper Danube – seem to be the Blätterhöhle individuals based on their position in the PCA, and their reasonable use for both potential admixture events involving Corded Ware or Yamnaya. I combined that Germany MN population in two-way admixture models including different potential Steppe-related sources:

Now, once we can discard Proto-Corded Ware and Corded Ware Poland, we should be able to compare closer populations to the target side by side. For that, I used BK_HUN_Sfu3 instead of Yamanya Samara – a priori a more reasonable comparison to the roughly coeval late Corded Ware group from Germany – following an empirically-proven strategy of adding reference populations, as explained here:
While the currently available statistical tools don’t seem to be ideal to clearly distinguish among Steppe-related origins, it certainly looks like adding outgroups increases the likelihood of finding the closest source of Bell Beakers – logically expected to be Yamnaya Hungary – even when the specific target group might be already admixed with peoples of Corded Ware ancestry.
At some point, which in this case was when adding the 19th outgroup, it seemed as though adding more references would almost invariably “worsen” the results, by obtaining higher p-values, higher SD or infeasible models, regardless of whether the added population was closer (EEF-, WHG-, EHG- or CHG related) or more distant (Asia, Africa). This process quickly becomes a boring game of finding a needle in the haystack, and not a very interesting one at that. One could assume that the general trend of higher p-values for Yamnaya compared to Corded Ware in these models would be interesting from a probabilistic point of view (since, experimentally, the highest p-value corresponds in ca. 50% of cases to the best qpAdm model), but this line of reasoning enters into the naïve, ruleless game of p-value ranking of ancestry magic I have warned about for a long time.
NOTE. Important caveats include the use of a late Bell Beaker sample from Hungary with an ancestry likely stemming from Moravia, thus showing Steppe ancestry probably partially admixed with Corded Ware, and likely a posteriori admixture with Yamnaya-related sources close to the Csepel region; and the use of a late Corded Ware family from Esperstedt including a high-Steppe outlier individual possibly related to direct admixture with Yamnaya vanguard groups. Not to mention that selecting the early Blätterhöhle MN is not ideal for any chronologically or regionally logical model: not only are reported high-HG groups surviving close to the Rhine, but there is also indirect evidence of survival of HG-rich groups in and around Hungary based on the ancestry found among Bell Beaker and Bronze Age samples of the region…

Upper Rhine BBC as origin of Lower Rhine BBC
Interestingly, the population that formed Beakers from the Netherlands and consequently the British Isles seems to be derived from an Upper Rhine Bell Beaker population close to Aes25, rather than from a Germany CWC-related source, despite the expected admixture events that must have happened downriver with Single Grave-related peoples:
Also relevant are the models for the high-Steppe male from the Althäuser burial, Alt_4, which suggest a slightly increased pull towards more Corded Ware-related ancestry, with one-way models of a high-Steppe-related source (Yamnaya, Corded Ware Germany, BK_HUN_Sfu3, or high-Steppe Poland_CWC) preferred over lots of infeasible models adding EEF(±WHG) populations, and where both CWC_DE and BK_HUN show similar results. A rather confusing trait that can only be explained (if the low coverage is not taken into account, by selecting the “AllSNPs: Yes” option) through its position in the PCA closest to all of these, in particular to BK_HUN_Sfu3, the Esperstedt outlier, or a Lech Valley Beaker individual (all in turn closest to the reported admixed Yamnaya Hungary EBA individual).
The ancestry of the Tauber Valley female, Alt_3, on the other hand, shows – as originally reported – a likely direct connection to the Balkans…
IV. Conclusion
We still don’t have the closest Steppe-related source of early Bell Beakers, and we won’t have it until the planned papers of the Reich Lab on the Hungarian Plains and South-Eastern Europe get published. Not much of a surprise expected there.
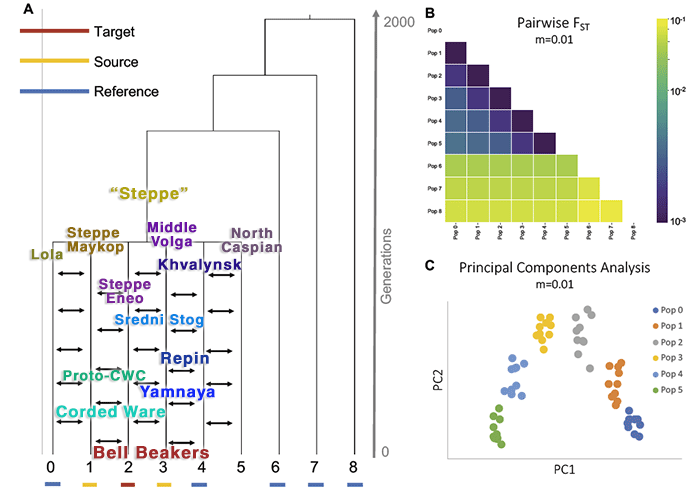
However, it is very likely that, even when we have a nice geographical and temporal transect including the Eneolithic and Bronze Age Steppe-related gene pool, distinguishing among Steppe-related populations will still be beyond the resolution of current analytical tools, based on the common mating networks of:
- The Steppe Maykop- and Eneolithic Steppe-like populations of the north Pontic area, including the late Sredni Stog and Proto-Corded Ware samples available to date.
- The EEF-admixed Repin (as shown by Afanasievo and Yamnaya) and likely relationship to Late Trypillia or a closely related population near the Don River, in turn connected to the (higher) EEF admixture found among Corded Ware individuals.
- The HG-admixed Poland/Baltic Corded Ware, potentially related to the north Pontic forest zone, given the HG-related shift of Ukraine Eneolithic samples, including the late Yamnaya/Catacomb outlier I5884.
- The Yamnaya vanguard settlers who must have admixed with local populations all over Central(-East) Europe, including Corded Ware groups, during the first half of the 3rd millennium BC.
- The Corded Ware-admixed East Bell Beakers that replaced late Corded Ware groups along the Upper Danube and to the north of it.
Luckily enough, this ship of Theseus-like paradox of Steppe ancestry, now also visible in CHG/Iran-related ancestry in the Near East, can be solved following straightforward cultural evolution models and the bottlenecks of Y-chromosome haplogroups that are known to happen during migrations, and Proto-Indo-Europeans seem to have expanded in two quickly successive waves of R1b-rich peoples, constantly reducing their Early Eneolithic haplogroup variability.
After all, a simplistically assessed similar “Steppe ancestry” component or cluster in a PCx vs. PCy plot of succeeding populations shouldn’t be that relevant, and cultural connections shouldn’t be made up based on a handful of continued “local” traits, or else researchers would still be speculating about Bell Beaker communities popping up out of nowhere continuing “local” populations from Iberia, or France, or the Netherlands, or Scandinavia, or Germany, or Moravia, or Poland… Oh, wait.
Related
- qpAdm best practices and common pitfalls
- Early arrival of Steppe ancestry in Switzerland
- Fully Steppe-like Proto-Corded Ware Late Trypillians
- “Steppe ancestry” step by step (2019): Mesolithic to Early Bronze Age Eurasia
- Bell Beakers and Mycenaeans from Yamnaya; Corded Ware from the forest steppe
- On the Ukraine Eneolithic outlier I6561 from Alexandria
- Yamnaya replaced Europeans, but admixed heavily as they spread to Asia
- East Bell Beakers, an in situ admixture of Yamna settlers and GAC-like groups in Hungary
- Trypillia and Greece Neolithic outliers: the smoking gun of Proto-Anatolian migrations?
- Yamnaya ancestry: mapping the Proto-Indo-European expansions
- East Bell Beakers, an in situ admixture of Yamna settlers and GAC-like groups in Hungary